| .Жизнь и Разум | |||
| . | |||
"Все собаки - сукины дети"
Уильям Филдс
"Порода сильнее пастбища"
Джордж Элиот |
| ДНК, гены, белки |
| Данная тема в мобильной версии изложена со значительными сокращениями. Не раскрыты вопросы генетического кода, копирования генов матричными РНК, синтеза белков и другие (перекрывающиеся гены, рибосомные РНК, хромосомы). Оставлена без рассмотрения "загадка" первичных белков и генов. С указанными изъятиями полное понимание биохимической сущности жизни, конечно, не возможно. В должной мере и в полном объеме все эти вопросы изложены в одноименной теме на стационарной версии сайта. В таком ее виде самостоятельное изучение всех ключевых вопросов химии жизни (в достаточной для понимания степени) вполне возможно. Но даже укороченная версия темы даст пользователю достаточно хорошее представление о молекулярном устройстве жизни. |
Молекулярная биология основана физиками-ядерщиками в 30-е - 50-е годы XX столетия. "Чистые" биологи в те времена ограничивали свою глубину познания жизни еще только уровнем оптических микроскопов и многолетних наблюдений. Важнейшим итогом этого этапа работ было открытие ДНК, РНК и генов. (Объяснение, что это такое, будет приведено ниже). Современные популяризаторы молекулярной биологии обычно объясняют что такое ДНК, как она устроена и "работает", но, как правило, не знают (или делают вид, что не знают), откуда она (точнее, они, разные ДНК) взялась(лись). Или же не придают этому вопросу должного внимания. А, может, просто не умеют сделать это достаточно ясным образом, простыми, понятными всем словами. Этим обстоятельством с удовольствием пользуются оппоненты теории самозарождения жизни. Из публикации в публикацию перекочевывает рассчитанная кем-то из них величина вероятности случайного самопроизвольного расположения атомов, совпадающая с их фактическим расположением в ДНК. Называются величины порядка 10 в минус 120-й степени и тому подобные ничтожные значения, для реализации которых, якобы, потребовалось бы число попыток большее, чем количество атомов во всей Вселенной. (Саму ДНК, ее строение и самостоятельную репликацию - деление на две части с образованием из них двух точных копий исходной - в наше время уже никто не подвергает сомнению). Судя по приведенному выше результату расчета, он мог быть получен только исходя из предположения, что все несколько десятков (или сот) миллионов атомов, входящих в состав крупной ДНК, одновременно и независимо друг от друга подошли из окружающего пространства к требуемым позициям и заняли их. Автор данной статьи глубоко убежден в бессмысленности такой методики расчета и о подобном способе самообразования ДНК. В природе все более сложное складывается из более простого, поэтапно, шаг за шагом. Поэтому данный сайт подходит к вопросу возникновения, устройства и "работы" ДНК прямо-таки диаметрально противоположным образом. Прямыми и не такими уж и сложными логическими соображениями здесь отслеживается последовательность образования сравнительно простых органических молекул из входящих в их состав атомов (см. предыдущую главу), а затем более сложных молекул из более простых. И так - вплоть до РНК, ДНК, генов и белков. Причем, это никакая не версия, а прямые (хотя и умозрительные) результаты наблюдения за последовательными физико-химическими преобразованиями в природе. Такой анализ основывается автором на общеизвестных и общепризнанных на сегодня исходных сведениях. Некоторые упрощения (а, возможно, и упущения) при описании "технологии" самозарождения и последующего существования ДНК и других компонентов живой материи допущены автором большей частью сознательно. И как непрофессионалом, излагающим материал для таких же непрофессионалов. Главное для всех нас - понять суть дела и прийти к правильным выводам. При дальнейшем ознакомлении с материалом данной статьи (как и предыдущей) надо понимать, что плоские (двухмерные) структурные формулы молекул или их фрагментов на листе бумаги или экране монитора предназначены только для схематической передачи основных связей между их ближайшими атомами. Реально эти молекулы имеют сложное пространственное строение, которое и определяет окончательно все взаимодействия между разными частями молекул и взаимодействия между разными молекулами. Передать графически такое строение всех молекул крайне сложно, и такая задача автором сайта, в принципе, не ставилась. С подобными проблемами мы все уже сталкивались при изучении плоских географических карт, а кое-кто - еще и при изучении карт звездного неба. Из этих примеров становится ясно, что кое-что для понимания объемного мира все-таки дают даже и его схематические плоские модели. Но некоторые молекулы представлены и в объемном виде, для наглядности, лучшего понимания и большей привлекательности всей данной статьи. Они выстроены автором с помощью специальных химических программ, в достаточном количестве имеющимися на просторах Интернета).
|
При наличии даже небольшого (по числу видов) количества биогенных химических элементов, а также условий для быстрого протекания различных химических реакций (в идеале - воды в жидкой фазе), в природе очень быстро создаются десятки и сотни тысяч самых различных химических соединений, разного состава и разной структуры. Подавляющее большинство из них так и остаются никем не востребованными растворами, осадками или мертвыми породами. Но некоторые из этих соединений, с виду - мало чем, в принципе, от других отличающиеся, впоследствии обнаруживают способность создавать удивительные комплексные соединения уже и между собой.
В данном конкретном случае такая способность вызвана двумя свободными валентностями, по одной на двух атомах кислорода. При наличии в окружающей среде каких либо атомов, они быстро присоединяются к свободным отросткам радикала. Обычно это бывают атомы водорода (гидрогена), наиболее распространенного во Вселенной химического элемента. И вместе с этими атомами этот радикал обычно и изображают. (Хотя после этого он, по существу, перестает быть радикалом в полном смысле этого слова). Большинство исследователей относят этот радикал к одному из "первокирпичиков" жизни. Невозможно спорить о важности этой структуры для возникновения и последующего развития жизни.
Во-первых, в начале формирования звездно-планетных систем (в т.ч., Солнечной системы и планеты Земля) существовал не только "дефицит" благоприятных условий (о них тогда просто не могло быть и речи), но и дефицит требуемых материалов (атомов), которые в некотором ограниченном пространстве могли быть наперечет, буквально поштучно. Поэтому на таких ранних стадиях развития природы более вероятным было появление как раз более простых соединений. В принципе, теоретически, некоторые реакции возможны даже и в открытом космосе, но там они крайне маловероятны, а если и случаются, то из-за большой пространственной разобщенности отдельных атомов протекают крайне медленно и могут нами вообще не учитываться. Во-вторых, современными методами исследований показано и доказано, что во всех главных жизнеобразующих молекулах их фосфорная оконечность имеет три (а не четыре) атома кислорода.
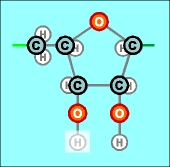
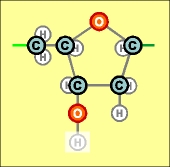
Как уже показано в предыдущей статье (что, впрочем, и без того достаточно очевидно), подобные соединения (или их фрагменты) легко образуются самостоятельно, без какого-либо внешнего вмешательства. Лишь бы были исходные материалы, в данном случае - атомы фосфора, кислорода и водорода и углерода. Другой относительно простой совокупностью атомов, легко соединяющихся в молекулы, являются карбонатные или углеродные (С) радикалы. В ходе развития природы и формирования жизни они стали центральными (в т.ч., и чисто геометрически) звеньями любой биологической молекулы. Их таких существует всего два вида, да и то, едва различимых между собой (в правом блоке - на один атом кислорода меньше):
Эти радикалы содержат пятизвенные кольца, называемые пентозами. (Бледным оттенком показаны атомы Н, которые могут замещаться в химических реакциях, например, при соединении пентоз с рассмотренными ранее фосфатными радикалами). Есть ли в этих структурах что-нибудь особо сложное, не поддающееся пониманию и силе воображения? Хотя бы в том смысле, что и эти химические соединения безо всяких затруднений могли образоваться самостоятельно? Конечно же, нет, представить такое никакой трудности не составляет. Подобные структурные самообразования при наличии исходных материалов в достаточном количестве и на близком расстоянии друг от друга не только возможны, но и просто неизбежны. Как показано в предыдущей главе, нет ничего необычного и в самостоятельном образовании немного более сложных кольцевых структур, содержащих атомы азота. Главное, как всегда, - наличие достаточного количества "строительных" материалов. Которые, к тому же, в качестве первых шагов без каких-либо затруднений могут образовывать соединения типа СН4 и NН3. Первый, самый простой (из тех, которые участвуют в формировании биологических молекул) тип азотистого (или по другому - нитратного) радикала, широко распространенный в природе, представляет собой такую двойную пару, (два его варианта, аналогично случаю с С_r и C_d):
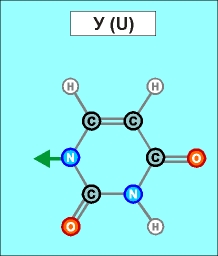
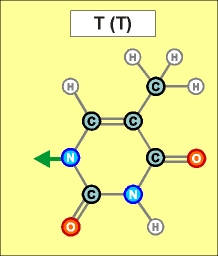
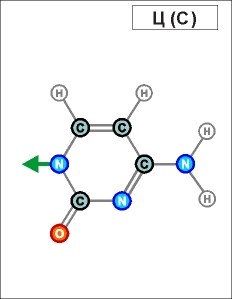
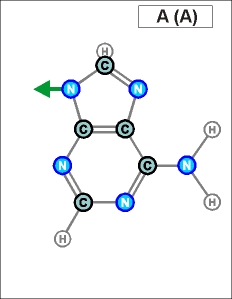
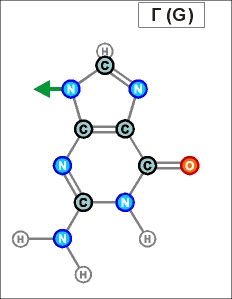
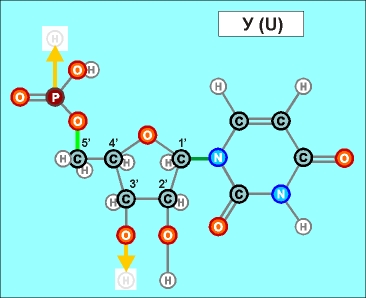
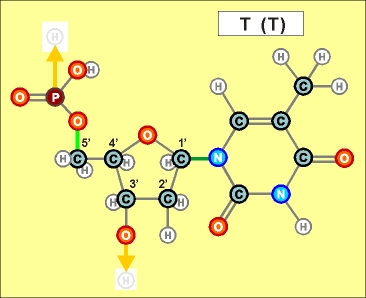
Урацил и тимин настолько похожи друг на друга (как и их соединения), что даже в таблице генетического кода (см.конец данной темы) вместо последовательностей Т (как раз и описывающих гены) часто указывают соединения группы У (U), хотя они относятся уже не к самим генам, а только к их "отпечаткам" - транспортным РНК. Существуют и другие природные азотистые радикалы. Например, такие (здесь выбраны только те их них, которые входят в формулы биологической химии, в состав живых клеток и их составляющих):
Белый цвет фона трех последних рисунков означает, что данные радикалы являются универсальными и могут входить в состав любых биологических молекул. (О важных различиях радикалов, изображенных на голубом и желтом фоне, будет рассказано далее). Все пять азотистых радикалов являются основаниями будущих нуклеотидов. Что это такое, см. уже в следующем абзаце.
|
Все представленные выше радикалы, свободные или временно "подгруженные" одним-двумя простейшими атомами, вступают в реакции при всякой встрече с "подходящими", более весомыми соединениями (по типу реакций замещения), в том числе, и в реакции между собой, образуя в конечном итоге нуклеотиды.
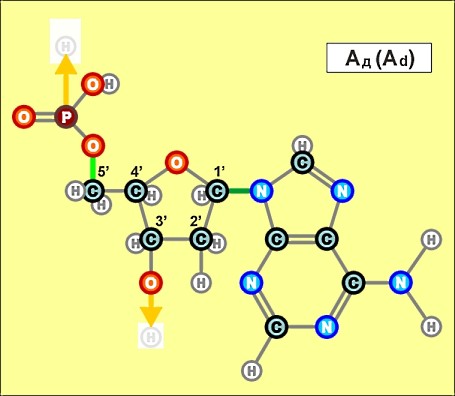
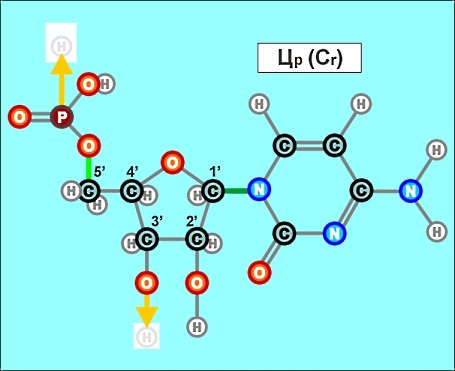
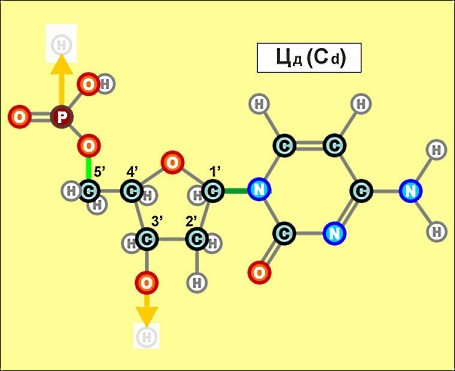
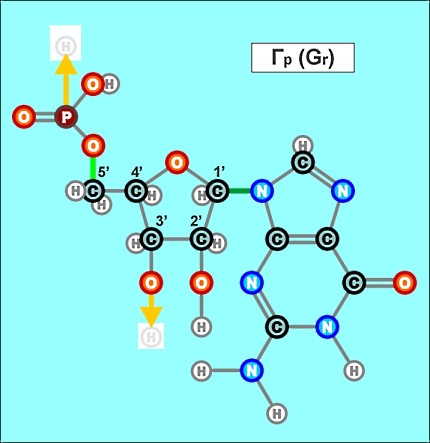
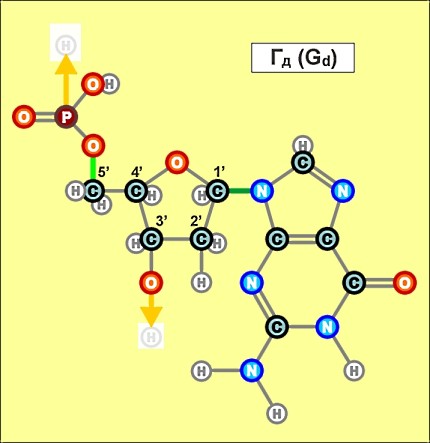
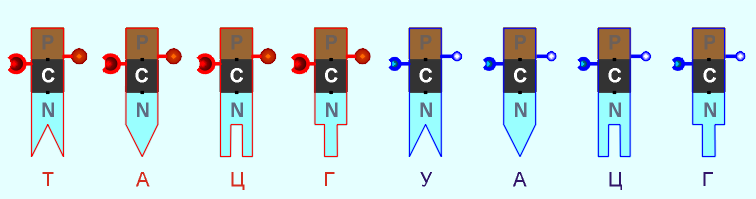
Пришло время указать, что все радикалы (и их соединения), представленные на рисунках с голубым фоном (имеющие по дополнительному атому кислорода/оксигена у второго справа атома углерода/карбона в пятиугольниках/пентозах), вступают в реакции и образуют устойчивые связи только между собой и с универсальными основаниями, показанными на рисунках с белым фоном. И они сами, и образованные ими нуклеотдиды относятся к т.н. соединениям урацилового ряда - по названию самого первого (и самого относительно простого) азотистого основания этой группы - урацила (сокращенно У или U). Аналогично, все радикалы и их соединения, представленные на этом сайте на рисунках с желтым (и белым) фоном, вступают в устойчивые постоянные связи только в пределах своей группы. В их карбоновой сердцевине/пентазе отсутствует один "необязательный" атом кислорода, из-за чего сам этот радикал и все его последующие соединения в своих полных наименованиях и имеют начало "дезокси" ("обескисленные"). Из-за этой "мелочи" существенно меняется структура электронного облака в нижней части любого нуклеотида этого ряда, из-за чего становятся невозможными их прочные соединения с продуктами противоположного, урацилового ряда. Чтобы лучше понимать, как идут дальнейшие процессы, не прибегая к слишком длинным записям, нуклеотиды условно обозначаются первыми буквами образовавших их оснований У(U), Ц(C), А(A), Г(G), Т(T), Ц(C), А(A), Г(G). При этом иногда их выделяют цветом или индексами r или d (чтобы отнести их к тому или иному ряду, особенно, когда названия совпадают). Основополагающим нуклеотидом этого ряда является тимин (Т). По первой букве приставки "дезокси" все они (кроме самого тимина) и их карбонатный радикал обозначаются, в случае необходимости, индексом d
Как же выглядят эти легендарные нуклеотиды? Опять ничего особенно сложного, этого всего лишь соединения рассмотренных нами ранее радикалов, по три всякий раз:
Здесь необходимо еще раз отметить, что данные плоские изображения далеко не в полной мере отражают внешний вид и особенности пространственного строения молекул, из которых, в значительной мере, вытекает и ряд их важных свойств. В первом приближении можно считать что левая (фосфатная) и правая (азотистая/нитратная) часть каждого нуклеотида лежит примерно в одной плоскости, а центральная часть (карбонатные пентозы) - практически в перпендикулярной. Это объясняется, в первую очередь, большой концентрацией атомов кислорода в соответствующих частях структур и сильным взаимным отталкиванием электронных облаков вокруг них. Для последующего перехода к построению и рассмотрению устройства более сложных, уже однозначно биологических объектов (РНК и ДНК), имеет смысл графически представить каждый нуклеотид в предельно упрощенном, условном схематическом виде, например, в таком:
Замена правой части формулы на другие существующие в природе азотистые радикалы и подбор соответствующих С-радикалов (с дополнительным атомом кислорода возле условно 2-го атома углерода или без него) создают нуклеотиды следующих видов:
Схематически они могут изображаться, например, так:
Еще одна пара возможных сочетаний радикалов:
Схематически эту пару можно изобразить так:
И, наконец, последняя пара возможных сочетаний (Рис. 12.1, 12.2):
Обозначим их схематически так:
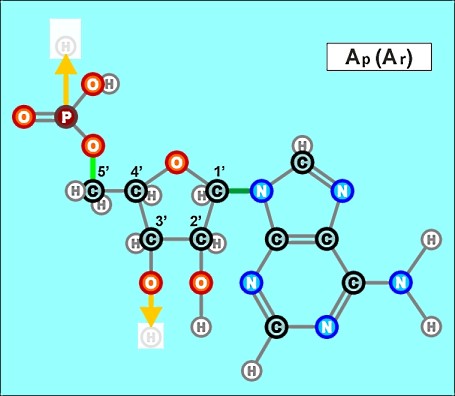
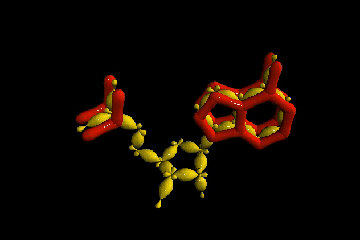
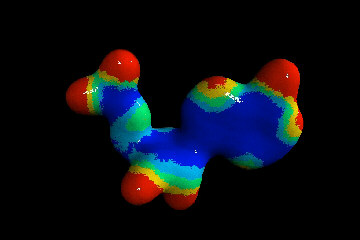
В очередной раз вспомним, что этими условиями являются только наличие материалов (нескольких типов биогенных атомов) и среды, в которой они и их соединения могут взаимодействовать (в идеале - воды). Чтобы получить более правильное представление о пространственном устройстве молекл нуклеотидов, а также для переключения внимания читателей из сферы умственной деятельности в область эстетического(!) восприятия учебного материала, ознакомимся с несколькими моментальными "снимками", сделанными в специализированных химических программах построения объемных (да еще и движущихся!) моделей молекулярных соединений. В качестве примера выбран нуклеотид аденин А_r (А_р). Разные его модели выстроены по разным методикам и с учетом разных приоритетов. Ниже представлена лишь незначительная часть результатов при различных подходах к их построениям. Атомы водорода, размещенные вплотную к тяжелым ядрам, образующим скелеты молекул, на рисунках условно не показаны. Цвета остальных атомов в моделях, выстроенных в разных компьютерных программах, совпадают не всегда, но принципиально это ничего не меняет.
Движущаяся объемная модель позволяет рассмотреть молекулу аденина (А р) с разных сторон:
Итак, как показано выше, природа создала нуклеотиды. Что происходило с ними дальше? Точнее, происходило, происходит и будеть происходить впредь! (В чем мы и убедимся в следующей подтеме).
|
||||||||||||||||||||||||||||||||||||||||||||||||||||
На следующем этапе самопроизвольных преобразований нуклеотиды урацилового ряда выстраиваются в цепочки так называемых РНК - рибонуклеиновых кислот. (Именно по первой букве этого слова в латинском начертании все радикалы и комплексы урацилового ряда и имеют индекс r . Кроме самоочевидного урацила.)
Нуклеотиды противоположного семейства ("обескисленных" радикалов и нейтральных универсальных азотистых оснований) могут выстраиваться (и постоянно успешно делают это) в продольные нити будущих ДНК - дезокси-рибонуклеиновых кислот. (Это длинное слово здесь написано через дефис только для удобства правильного его первого прочтения. По первой букве этих кислот все исходные радикалы этого ряда имеют индекс d . Кроме стоящего особняком тимина.) На предыдущих схематических рисунках нуклеотидов бледным цветом были показаны атомы, в направлении которых каждый нуклеотид сильно поляризован, потому что с одной его стороны почти по одной прямой линии расположено сразу три атома кислорода, а с другой (после третьего атома углерода С 3' ) - только один. Имеющийся у всех нуклеотидов уринового ряда еще один атом кислорода, возле атома С 2' карбоновой пентозы, как раз этим дополнительным атомом углерода и нейтрализован (как другая возможная точка поляризации). Более того, "лишний" атом кислорода только немного перераспределяет напряженность электрического поля у атома С 3' , делая все продольные соединения нуклеотидов этого ряда немного слабее. Так или иначе, вертикальный (по рисункам) поляризованный участок любого нуклеотида (Н)-ОРОССС-(ОН), или, в другой форме записи ("радикальной"), -OPOCCCO- , является универсальным (и очень прочным!) продольным соединителем, действующим по принципу электростатического притяжения. К тому же, сблизившиеся за счет этого звенья укрепляют свое взаимодействие еще и за счет создания совместных (частично) электронных оболочек. Такое взаимодействие неизбежно приводит к естественной (хотя и вполне спонтанной) полимеризации нуклеотидов - их соединению в длинные вертикальные нити. Наличие большого количества свободных (одиночных) нуклеотидов в органических растворах (в т.ч., и в так называемом первичном биологическом бульоне) способствует этому процессу, а их дефицит тормозит его, но принципиально абсолютно ничего в нем не меняет. Нуклеотиды обоих семейств очень быстро выстаиваться в очень длинные цепи, причем, вначале (на стадии самообразования) - без какой-либо системы в этих цепях, например: ...УЦЦААУГГГЦЦУУАУГГГЦЦЦУАУАУАГГАЦЦАУУ...... и т.д. В цепях "дезокси"-типа вместо урацила У в ряды встраивается тимин Т. Длина такого рода цепей может достигать десятков или сотен тысяч нуклеотидов (у бактерий и простейших одноклеточных), а то и многих миллионов (или даже миллиардов - у высших биологических видов). Причиной высокой степени полимеризации (образования длинных нитей) является исключительно высокая прочность продольных соединителей.
При этом нуклеотиды урацилового ряда (обозначенные на предыдущих графических материалах оттенками синего цвета и индексом r), рано или поздно выстраиваются в нити (одиночные) рибо-нуклеиновой кислоты (РНК), а тиминового, то есть, "дезокси"-ряда нуклеотидов (обозначенных на всех предыдущих рисунках оттенками желтого и красного, а также индексом d), - в нити дезокси-рибо-нуклеиновой - ДНК. (Причем, во втором случае сразу происходит наращивание и второй, параллельной ее цепи, см. далее). Графически универсальный продольный соединитель можно условно изобразить, например, в виде шарового замка-защелки (подобного тем, которые используются в детских пластмассовых конструкторах):
В условном графическом представлении, для наглядности, это показано в виде шаровых соединителей разного диаметра и цвета. В последующих графических построениях продольные соединители не показываются для исключения чрезмерного загромождения рисунков, которое только мешало бы пониманию процессов. Но их наличие постоянно подразумевается по умолчанию. Цепи ДНК гораздо длиннее, чем РНК. Дополнительный отросток ОН, имеющийся в РНК, несколько затрудняет процесс полимеризации (за счет некоторого размывания и смещения электронного облака, см. движущуюся объемную модель выше), и делает продольные связи РНК менее прочными.
Со 100%-ной уверенностью ясна лишь причина ограниченности длины РНК для матричных РНК, копирующих информацию генов (см. ниже). Это строгая заданность структуры ее начала и конца копируемым ею геном. Дальнейшее рассмотрение будем проводить, начиная с ДНК. Ее роль в образовании и поддержании жизни, особенно, на первом этапе, все-таки, намного важнее. А главное, - изначально она формируется из более простых (хоть и не намного) составляющих. При очень длинных цепях, их подвижности и гибкости (из-за теплового фактора, наличия оболочки, то есть ограничителя длины прямолинейного расширения и др. причин), а также при постепенном снижении концентрации неохваченных указанным процессом нуклеотидов, цепи могут замыкаться в кольца, а иногда даже запутываться в узлы. Для этого только необходимо, чтобы конец цепи случайным образом достаточно приблизился к ее началу. Случай случаем, но при таких длинных и произвольно ориентируемых на каждой стадии образования цепях это вполне может происходить. Ничего сверхестественного в этом, конечно, нет. Замыкание цепи ДНК в обычное кольцо, без узлов, - еще и очень выгодное для ДНК состояние, как самое прочное. Вместе с тем, это, все-таки, не строго обязательное условие образования ДНК.
После замыкания в кольцо цепь приобретает особую прочность и устойчивость, так как в дальнейшем вся ее подвижность сводится к "утряске", то есть, занятию всеми ее фрагментами и отдельными атомами наиболее устойчивых в энергетическом смысле положений. При этом постепенно начинает формироваться и пространственная структура цепи. В реальной ситуации одновременно (или чуть позже) с процессом наращивания длины цепи идет еще и ее утолщение в 2 раза за счет формирования параллельной цепи, соответствующей (но не равной!) первой. Причиной этого является наличие в каждом нуклеотиде азотистого радикала (см. представленные выше рисунки), стремящегося воссоединиться с другим, подходящим ему (или, как принято говорить, комплементарным, ему). Что же это такое, комплементарность? Не надо пугаться этого якобы сложного слова. Его надо воспринимать как комбинацию более простых, привычных и приятных слов комплект и комплимент, что соответствует (случайно или специально - автору не известно) и сути новообразованного слова:
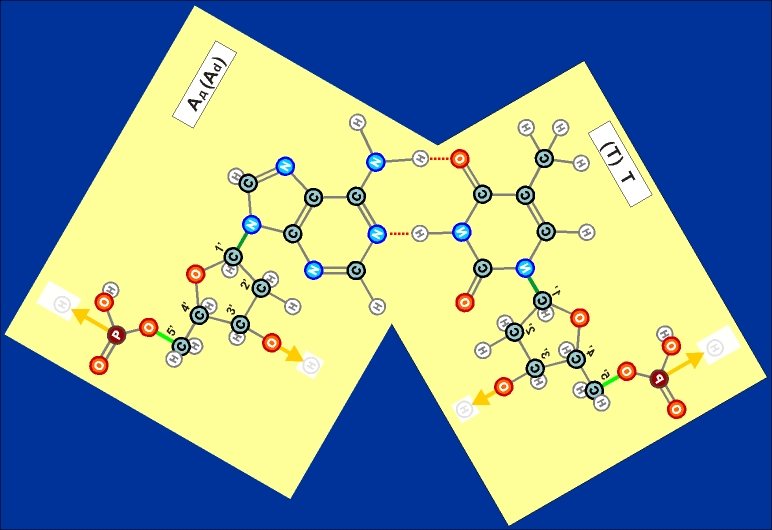
В чем заключается эта "подходящесть", станет ясно из следующего ее описания и поясняющих рисунков. Как показано выше, два из нуклеотидов (Т и Ц) содержат по одному азотистом кольцу , а два (А и Г) - по два кольца. Одно кольцо содержит и нуклеотид У. С точки зрения его строения он является близким аналогом Т, но из-за большей степени окисления своего карбонатного радикала может входить только в состав цепей РНК (как и РНК-овские варианты А, Г и Ц). Свободные нуклеотиды, оказывающиеся поблизости от одиночной (на первом этапе формирования) цепи ДНК, притягиваются к ней сначала дополнительными атомами азотистых колец таким образом, чтобы за их счет на исходных кольцах как бы образовывалось(лись) еще одно, а то и два новых, связывающих псевдо-кольца (см. рисунки 14.1 и 14.2, на которых они обозначены красным пунктиром). Псевдо, - потому что в "полноценное" (и окончательное) химическое соединение "чужие" азотистые кольца так и не вступают, а только притягиваются друг к другу по принципу комплементарности (см. выше). Такое сближение соответствует устойчивому состоянию образующейся при этом пространственной структуры их двух нуклеотидов.
Рис. 12.1 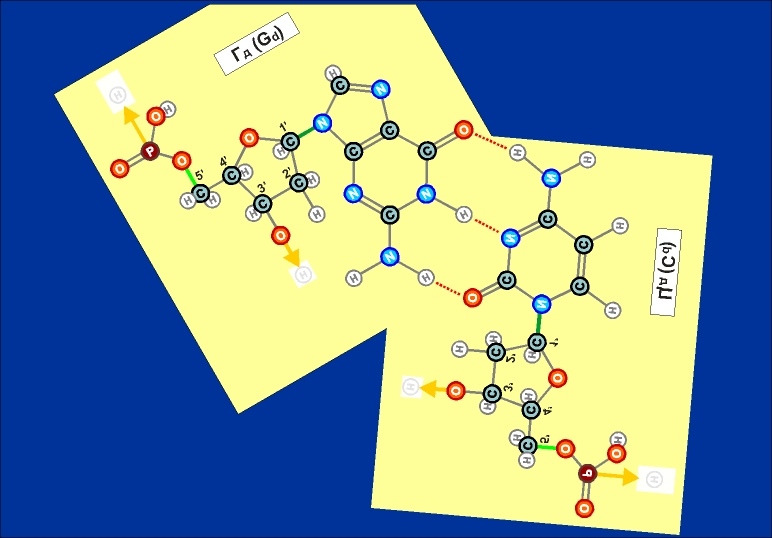 Рис. 12.2 Как видно из этих рисунков, из-за расположения и влияния "второстепенных", дополнительных атомов при "главных" кольцах, боковой захват возможен только между парами А - Т и Г - Ц (в РНК в первой паре вместо Т взаимодействует его аналог У). Это и есть иллюстрацией (и объяснением причины) комплементарности этих пар.
Из этих же рисунков видно, что соединение А - Т (У - Т), через две линии взаимодействия (и одно дополнительно возникающее кольцо) значительно более слабое, чем соединение Г - Ц, через три линии взаимодействия (и через два дополнительных кольца). Таким образом, допустимыми боковыми соединениями нуклеотидов (для их попарного существования - главными) возможны только следующие (Таблица комплементарности):
Очень важным является то, что к любому нуклеотиду семейства ДНК (на рис. 13 - красные) могут временно присоединяться не только "свои" комплементарные полу-пары, но и аналогичные нуклеотиды семейства РНК (У, А, Ц, Г), обозначенные на рис.13 справа, синим цветом. Те, которые отвечают принципу комплементарности. Например, к "красному" Т вполне может присоединиться "синий" А; их соединители взаимно подходят друг другу. Однако такие соединения бывают только временными (из-за относительно слабой боковой связи между ними, которая, в свою очередь, объясняется хоть и не очень значительным, но вполне конкретным отличием в строении "красных" и "синих" нуклеотидов). Но свою жизненно важную роль они еще сыграют (например, при копировании участков цепи ДНК). После бокового присоединения любого нуклеотида к формируемой цепи он немедленно соединяется через свой универсальный продольный соединитель еще и со своим соседом, присоединившимся к цепи на мгновенье раньше, формируя таким образом еще одну продольную цепь (ветвь), параллельную первой:
В пространстве этот процесс (его небольшая часть) может выглядеть примерно так:
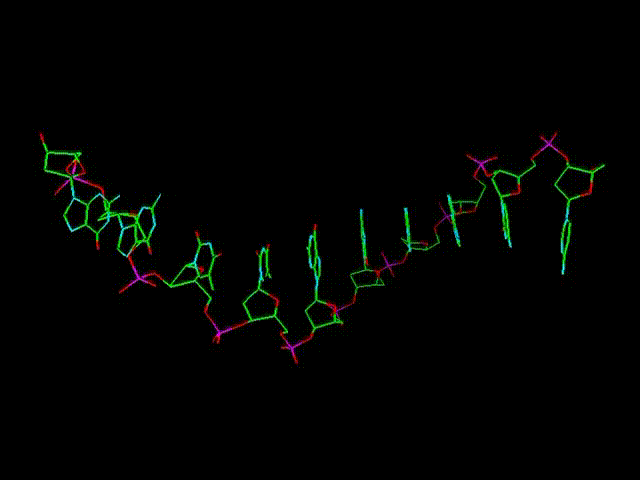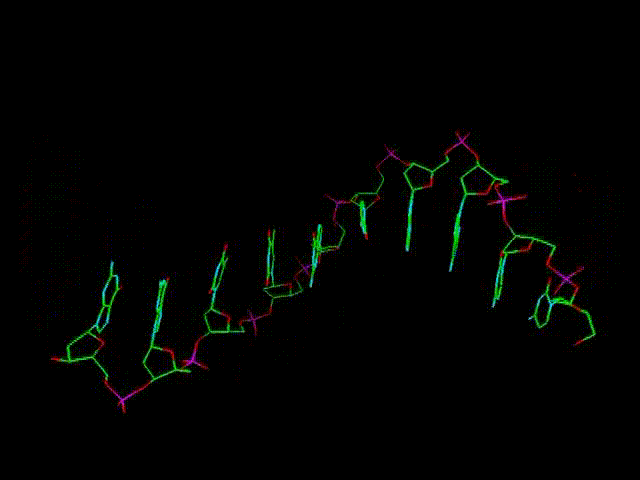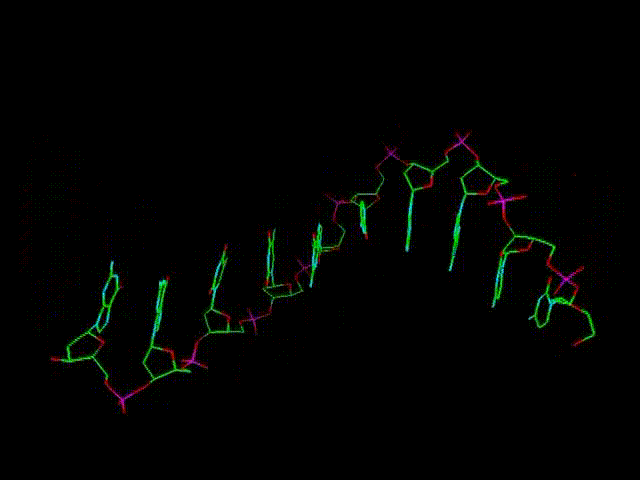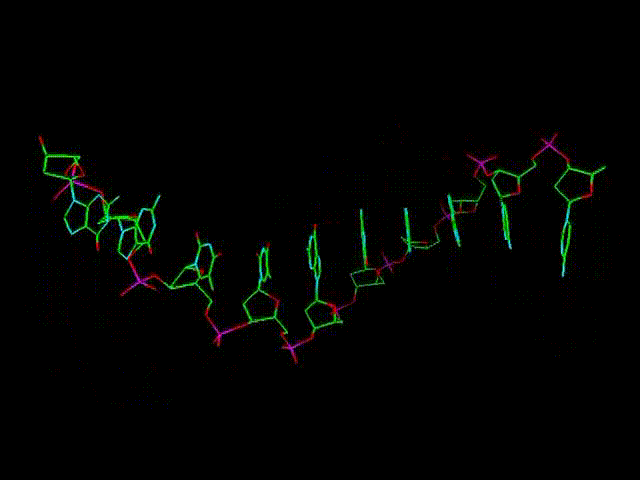
По мере полного заполнения всех боковых "вакансий" образуется двойная цепь, туго закрученная в спираль, см. рисунок в самом начале темы. Это и есть молекула ДНК. Еще одной особенностью ДНК является противоположное по поляризации (ориентации) направление ветвей (после завершения ее построения). Если одна из них как бы идет вправо (или в любом другом направлении), то другая - влево (или в любом другом направлении, противоположном первому).
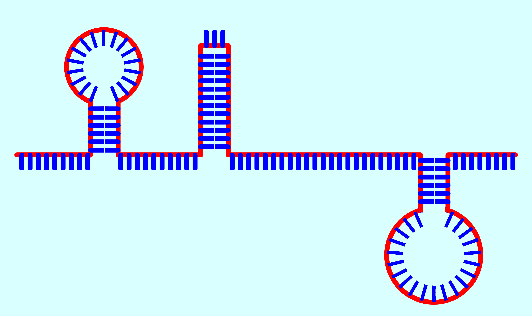
Зато первичные (не матричные) РНК имеют интересное свойство частично складываться в отростки ("шпильки" или петли), если на небольшом расстоянии цепи РНК попарно имеются отрезки, комплементарные друг другу, которые и соединяются между собой (синим цветом на рис. 14 показаны нуклеотиды, красным - образующаяся продольная нить):
Пространственная спираль ДНК - следствие не плоской и не абсолютно симметричной (при виде с условного "верха") структуры исходных нуклеотидов, а также создания продольных связей через разные атомы пентаз, от т.н. 5-го атома углерода нижней пентазы (фактически - четвертого, через дополнительный 5-й) к 3-му следующей верхней. Тем не менее, образующаяся спиралевидная молекула получает дополнительную устойчивость еще и за счет этого. Двойной характер цепи ДНК - следствие параллельно идущего продольного и бокового сцепления подключающихся к первичной цепи свободных нуклеотидов (которых в любом биологическом растворе всегда предостаточно). Замкнутые в кольца ДНК способны закручиваться и в "сверхспирали", что позволяет им быть прочными и очень компактными. Иллюстрацией сверхспирали может служить многократно закрученная бельевая веревка или шнур пылесоса, которые способны создавать произвольные самозакручивающиеся петли где-то в их середине.
|
Из описанного выше процесса самообразования двойной спирали ДНК, состоящей из случайной последовательности нуклеотидов, вытекает предположение об огромной, чуть ли не бесконечной многовариантности таких первичных ДНК. А такого, мол, нет и не может быть. Не наблюдаем мы бесконечного количества видов ДНК (и образовавшихся на их основе биологических видов), даже при помощи самых современных методов. На самом деле, практически наблюдаем. Несколько сот тысяч биологических видов на Земле, включая растительный мир, животных всех видов, птиц, насекомых, рыб, моллюсков, бактерий, микробов и вирусов - это очень большая многовариантность. А ведь это - только ничтожная часть из всей первичной многовариантности случайно образующихся ДНК из-за далеко не обязательной способности абсолютно всех их к формированию живых организмов. Или, тем более, способных бороться за выживание в бесконечно конкурентной среде. И, тем более, побеждающих в этой борьбе. Только последние и есть то, что мы видим сейчас вокруг себя. Подавляющее же большинство первичных ДНК достаточно короткие и относительно примитивные, способные формировать только ничтожные комочки аминокислот - наипримитивнейшие белки (см. далее), которые пригодны только для того, чтобы пойти в пищу более сложным и совершенным. Кстати, ничто не мешает формироваться таким первичным (не наследственным) ДНК и сейчас. Но они тоже в своем абсолютном большинстве просто поедаются всеми, даже простейшими из уже сформировавшихся на сегодня видами, владеющими биосферой Земли. Но некоторые (единицы), все-таки, и формируются, и выживают. В форме простейших новых вирусов, агрессивных и вполне способных постоять за себя. Длина их ДНК составляет "всего" несколько тысяч нуклеотидов.
|
После описанного выше завершения формирования любой из первичных ДНК обе ее продольные цепи закручиваются и стягиваются настолько сильно и прочно, что это приводит к разрыву некоторого участка поперечных связей. (Это немного напоминает разрывы нитей полотенца при его чрезмерном выкручивании после стирки). Правда, многие считают причиной такого разрыва не чрезмерное закручивание, а как раз наоборот, раскручивание участка спирали (например, под действием тепла) и ослабевание поперечных связей (по той же или еще какой-нибудь причине). Так или иначе, на некотором участке ДНК (для незамкнутых в кольцо учебных моделей - с краю) возникает разрыв поперечных связей и ее нити (цепи) несколько расходятся.
Рис.15 В образовавшийся разрыв тут же устремляются многочисленные свободные нуклеотиды, в изобилии содержащиеся в окружающей среде (точнее, уже в клетке). Они комплементарно (по принципу "подходящести", см. выше) присоединяются к "раскрытым" звеньям в каждой нити, и тут же сращиваются через универсальный продольный карбонатный (т.н. "сахарный") соединитель и между собой, образуя две новые двойные нити, ничем не отличающиеся от первичных, "родительских". Постепенно проходя через всю длину ДНК, этот процесс, в конце-концов, приводит к образованию двух новых ДНК, ничем не отличающихся от исходной.
Большими черными стрелками на Рис.15 показано направление физического разделения ветвей, а не упоминавшееся ранее направление их поляризации, которое противоположно друг другу, см. ниже, Рис.16.
|
Кажущаяся бесконечной и монотонной двойная цепь, визуально не слишком значительно оличающихся друг от друга нуклеотидов (особенно, в наружном сахарно-фосфатном остове), удивительным образом несет на себе всю информацию об абсолютно любом признаке будущего (или уже существующего) организма, для которого данная ДНК предназначена. Как же можно хотя бы представить себе такое? Да и возможно ли это вообще? Оказывается, да. Несмотря на кажущуюся бессмысленность во взаимном расположении нуклеотидов в каждой цепи, оно то, это расположение, и несет всю требуемую информацию. Оказывается, что лишь последовательности нуклеотидов ТАЦ или ЦАЦ (это т.н. стартовые кодоны) могут являться началом формирования на них новых биологических молекул (позже увидим, что это РНК), являющихся и своеобразным сигналом начала "записи" информации об очередном признаке биологического вида - так называемого гена.
Стартовыми последовательностями могут быть только триады ТАЦ или ЦАЦ. Далее они могут входить и в состав центральной части любого гена, по ходу его формирования (как и наблюдения или записи исследователем), и там они, конечно, уже предстают в качестве обычных триад, а не стартовых.
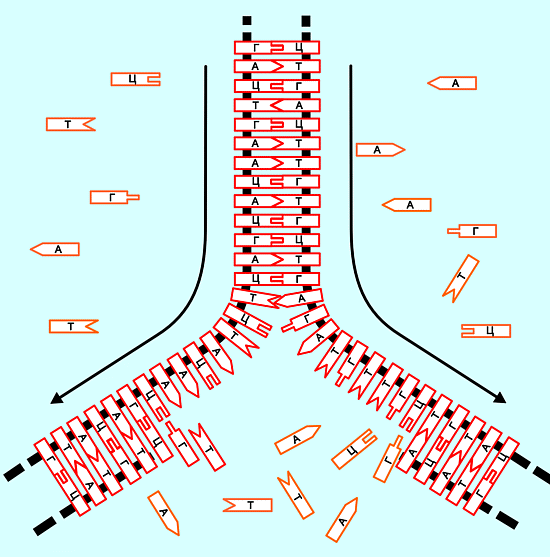
Существенным является то, что финишные последовательности должны входить именно в состав триад, разбитых от начала гена, а не быть элементами смежных триад. Поясняющий эту мысль пример (пробелы в записи - условные, для наглядности разбиения гена на триады): ...АГГЦАТАТАТАЦ ЦАТ ГГЦ ЦЦТ ... ТАГ ЦАГ АТА ... ГГГ АТЦГЦАЦЦГААЦ..., Выделенная красным цветом последовательность не является терминирующей, так как ее элементы входят в разные, хотя и смежные триады. Причины "старта" и "финиша" гена определяются особенностями последующего формирования белков организма из аминокислот (с помошью копирующих и транспортных РНК), последовательность которых в полной мере определяется триадами гена. При бессистемных, "не подходящих" начальных последовательностях нуклеотидов в ДНК, химия процесса просто не начинается. Подобно строительству высотного дома без фундамента. Или открыванию замка без подходящего ключа. Или приготовлению горячей пищи без источника тепла. В дальнейшем он идет непрерывно, практически независимо от состава последующих триад (толпу, врывающуюся в только что открытый магазин, музей или концертный зал, остановить почти невозможно). Обратная ситуация - в конце гена, при перемещении "считывающего устройства" (молекулы РНК) в зону одной из трех терминирующих триад. На основе их последовательностей сформировать никакой новый белок организму уже не удается (потому что в химии не все соединяется с чем попало). И их синтез прекращается. Строительная аналогия: монтажникам высотки подняли элементы конструкции крыши дома. После них дальнейшее возведение стен уже невозможно. Кулинарная аналогия: приготовление блюда заканчивается с использованием последнего подходящего ингридиента. Если дальше кто-то предложит положить в суп кирпич, хозяйка с негодованием отметет такое предложение. На следующем рисунке (см. ниже) приведена иллюстрация к понятию гена (и даже не одного), а также копирования с одного из них (с помощью РНК 1) "рецептов" (признаков) для начала строительства организма (Белок 1). Подробности - на стационарной версии сайта.
Рис. 16 В левой нижней части, на участке разделения цепей ДНК, условно выделенным зеленым цветом показано начало Гена 1 (триада ТАЦ, порядок рассмотрения - снизу вверх), желтым цветом - триады "тела" гена (их реальная длина - от нескольких десятков до нескольких десятков тысяч триад), а темно-серым - триада АТЦ конца гена. Последующие нуклеотиды являются нейтральным промежутком, "пробелом" между генами, именуемым промотором. Она будет безынтересной для любого "снятия отпечатка", пока в цепи не попадется очередная "стартовая" группа, подходщая для создания белка другого типа. В смежной ветви ДНК ориентация нуклеотидов противоположная (на рисунке - справа и сверху вниз). В ней могут оказаться абсолютно иные гены, независимые от генов первой рассмотренной ветви (показано начало гена 2). Но такие же важные для формируемого организма. Как же из случайной последовательности нуклеотидов в ДНК получается что-то толковое? Если случайно эта последовательность позволит (через РНК) свершиться всем полезным биохимическим процессам, и если необходимые для жизни белки (и органы из них!) будут сформированы, то создаваемый таким образом организм выживает и передает наследственную информацию своим будущим потомкам о правильности своей ДНК (и передает как раз через нее!) Из всех случайных последовательностей нуклеотидов, составляющих набор генов, закономерно, по наследству, сохраняются и передаются только уникально редко встречающиеся удачные. Это и есть первый шаг жизни.
|
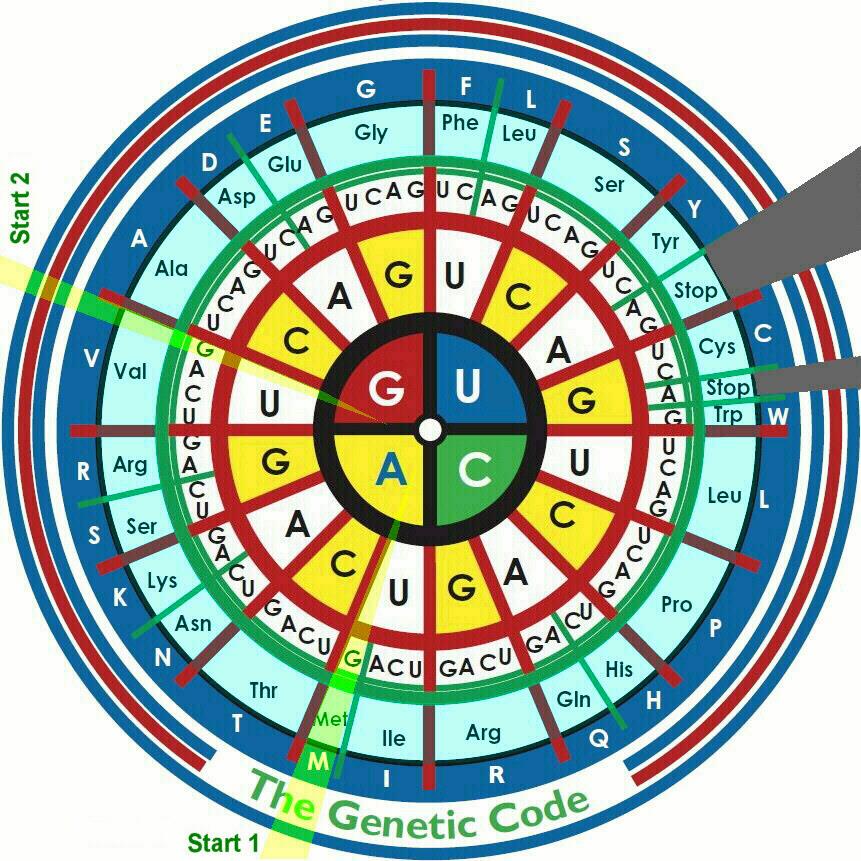
Генетическим кодом природы является соответствие формируемых организмами белков исходной информации о них, заложенной в генах ДНК. Для исследователя - это просто алфавит живой природы, своеобразная система условных обозначений, изложенная в виде таблиц или диаграмм. Таблица соответствий. И только.
А вот полный набор всех генетически передаваемых признаков каждого биологического вида и каждого его индивидуума - т.н. геном, конечно же, существует. Но разбор этого вопроса невероятно сложен для посетителей данного сайта. Как и для его автора-составителя. Остается только указать, что больше 99% генной информации у каждого представителя любого биологического вида полностью совпадает. Но те немногие доли процента, которые отличаются, как раз и делают каждого индивидуальностью, а у людей - еще и личностью. Вернемся, все таки, к генетическому коду. Фактически, в разгаданном виде, это просто результат исследований лаборантов. В каких случаях совмещаются те или иные аминокислоты, и что из этого получается. Хотя наследственная (даже и на самых ранних стадях) информация заложена в наборе генов каждой ДНК, ее воплощение в реальную жизнь проводят несколько специализированных типов РНК, то есть, соединений так называемого урацилового ряда. В связи с этим, таблицы соответствий триад генов получаемым в окончательном виде белкам ("генетический код") чаще записывают не в виде нуклеотидов d-ряда (как, по идее, должно быть и как все это излагалось на данном сайте ранее), а в виде сочетаний соответствующих (комплементарных) нуклеотидов урацилового (на данном сайте - "синего") ряда. Посмотрим еще раз на таблицу комплементарности (она уже была ранее):
Вспоминаем о стартовых и терминирующих (останавливающих) последовательностях нуклеотидов в генах (предыдущая глава) и находим соответствующие им комплементарно (попарно подходящие, хотя бы для временного соединения) последовательности РНК-нуклидов:
Эта таблица помогает понять методику построения и некоторые результаты в англоязычной таблице генетического кода, для удобства и красоты свернутой в круг. Читать последовательности r-нуклеотидов следует, начиная с центра круга. Результаты, в виде сокращенного названия получаемой аминокислоты - в четвертой от центра окружности (небесно-голубого цвета).
Более глубокие подробности в данной версии сайта не рассматриваются. * * * * * * * В этом месте мобильной версии сайта для экономии места исключен ряд важных вопросов (матричные, транспортные и рибосомные РНК, белки и их синтез хромосомы и др.) Но для понимания биохимической сущности жизни они, все таки, необходимы. Полное изложение этих материалов представлено на стационарной версии сайта, в теме с таким же названием (ДНК, гены, белки). Переход по ссылке.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| . | |||||||||
| . | . | ||||||||
| . | |||||||||
| . | |||||||||
| Следующая тема | |||||||||
| . | |||||||||